
Nitzschia A.H. Hassall; 1845; 435
Key references
Lange-Bertalot H. 1976. Eine Revision zur Taxonomie der Nitzschiae lanceolatae Grunow. Die "klassischen" bis 1930 beschreibenen Süßwasserarten Europas. Nova Hedwigia. 28(2-3): 253-308.
Lange-Bertalot H., Simonsen R. 1978. A taxonomic revision of the Nitzschiae lanceolatae Grunow. 2. European and related extra-European fresh water and brackish water taxa. Bacillaria. 1: 11-111.
Mann D.G., Trobajo R., Sato S., Li C., Witkowski A., Rimet F., Ashworth M.P., Hollands R.M., Theriot E.C. 2021. Ripe for reassessment: A synthesis of available molecular data for the speciose diatom family Bacillariaceae. Molecular Phylogenetics and Evolution. 158: 106985.
Morphology
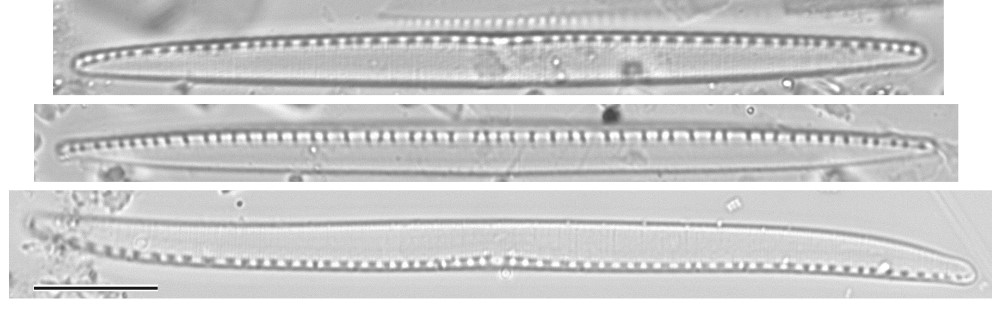
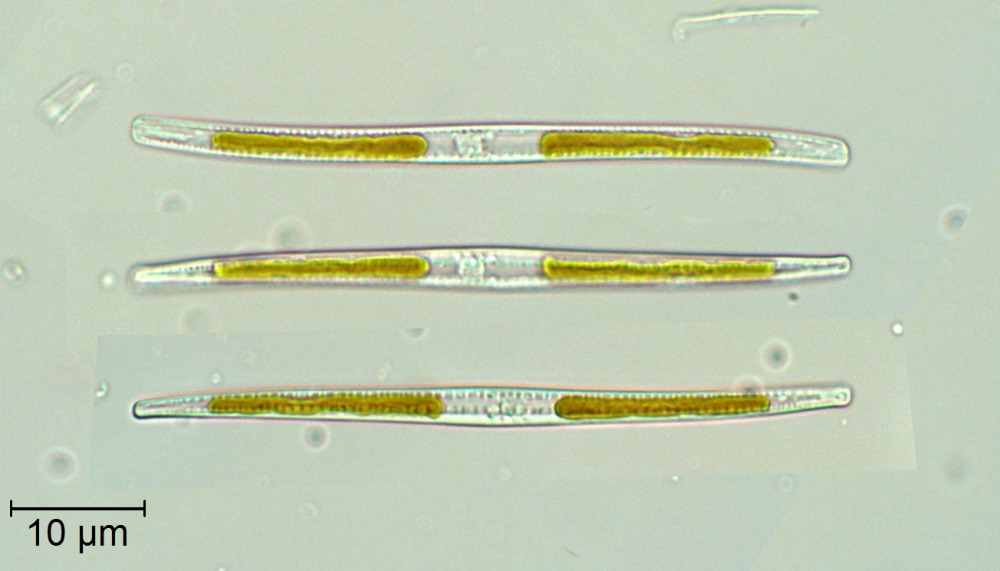
Cells often highly elongate, even spicular, but some species nearly isodiametric; valve outline highly variable: linear, lanceolate, elliptical, or sigmoid, with or without a central constriction; poles rounded or rostrate to capitate. In girdle view cells may or may not be constricted centrally, and may be straight or sigmoid (depending on the species, cells may be sigmoid in valve view or girdle view or both). Cells lie more frequently in valve view or girdle view, depending on the species.
Isovalvar, isopolar, more or less symmetrical in outline with respect to the apical axis but structurally asymmetrical because the raphe system is almost always placed off-centre.
The raphe systems of the two valves may lie on the same side of the cell ('hantzschioid symmetry') or on opposite sides ('nitzschioid symmetry'); species vary according to whether all cells are nitzschioid, all cells are hantzschioid, or nitzschioid and hantzschioid symmetries both occur, in ratios of either 1:1 or 1:2.
Striae usually uniseriate and parallel, with round or transapically elliptical areolae occluded by hymenes and sometimes by cribra too; the striae are rarely coarse, often delicate, sometimes so fine to be unresolvable in LM, even in moderate to large-celled species.
Raphe system fibulate, slightly to strongly eccentric, usually appearing on the edge of the valve but subcentral in a few species; except in a few cases (e.g. N. filiformis, N. obtusa), the raphe maintains the same position on the valve over its whole length. The raphe itself is almost always invisible, its presence being made apparent by the presence of fibulae. Central raphe endings present or absent (this is an important taxonomic character at the species level); where present, they are usually detectable in LM as a slight thickening of the wall of the raphe canal at the centre and/or by a wider separation of the central pair of fibulae. Fibulae rib-like, square or rectangular structures, usually small, but in a few species extending across part or all of the valve.
Generally two chloroplasts per cell, positioned ‘fore and aft’ (i.e. with one towards each pole); very occasionally >2 chloroplasts in some marine species; chloroplasts usually simple, appressed to one or other valve or to the girdle.
Cells usually solitary, more rarely forming stellate or chain-like colonies, or living in mucilage tubes.
Isovalvar, isopolar, more or less symmetrical in outline with respect to the apical axis but structurally asymmetrical because the raphe system is almost always placed off-centre.
The raphe systems of the two valves may lie on the same side of the cell ('hantzschioid symmetry') or on opposite sides ('nitzschioid symmetry'); species vary according to whether all cells are nitzschioid, all cells are hantzschioid, or nitzschioid and hantzschioid symmetries both occur, in ratios of either 1:1 or 1:2.
Striae usually uniseriate and parallel, with round or transapically elliptical areolae occluded by hymenes and sometimes by cribra too; the striae are rarely coarse, often delicate, sometimes so fine to be unresolvable in LM, even in moderate to large-celled species.
Raphe system fibulate, slightly to strongly eccentric, usually appearing on the edge of the valve but subcentral in a few species; except in a few cases (e.g. N. filiformis, N. obtusa), the raphe maintains the same position on the valve over its whole length. The raphe itself is almost always invisible, its presence being made apparent by the presence of fibulae. Central raphe endings present or absent (this is an important taxonomic character at the species level); where present, they are usually detectable in LM as a slight thickening of the wall of the raphe canal at the centre and/or by a wider separation of the central pair of fibulae. Fibulae rib-like, square or rectangular structures, usually small, but in a few species extending across part or all of the valve.
Generally two chloroplasts per cell, positioned ‘fore and aft’ (i.e. with one towards each pole); very occasionally >2 chloroplasts in some marine species; chloroplasts usually simple, appressed to one or other valve or to the girdle.
Cells usually solitary, more rarely forming stellate or chain-like colonies, or living in mucilage tubes.
Literature
References are given in chronological order.
Reference |
Citation |
|---|---|
| Hassall, A.H. 1845. A history of the British Freshwater Algae (including descriptions of the Diatomaceae and Desmidiaceae) with upwards of one hundred Plates. I. Text. Taylor, Walton, and Maberly, London. 462 pp.. | Morphology; Taxonomy; Description |
| Lange-Bertalot H. 1976. Eine Revision zur Taxonomie der Nitzschiae lanceolatae Grunow. Die "klassischen" bis 1930 beschreibenen Süßwasserarten Europas. Nova Hedwigia. 28(2-3): 253-308. | Morphology; Illustrations |
| Lange-Bertalot H., Simonsen R. 1978. A taxonomic revision of the Nitzschiae lanceolatae Grunow. 2. European and related extra-European fresh water and brackish water taxa. Bacillaria. 1: 11-111. | Morphology; Illustrations; Taxonomy |
| Krammer K., Lange-Bertalot H. 1988. Süßwasserflora Mitteleuropas. Bacillariophyceae. 2. Teil. Bacillariophyceae, Epithemiaceae, Surirellaceae. Eds. Ettl H., Gerloff J., Heynig H., Mollenhauer D., Gustav Fischer Verlag, Stuttgart. 596 pp | Morphology; Taxonomy; Illustrations |
| Trobajo R., Mann D.G., Chepurnov V.A., Clavero E., Cox E.J. 2006. Taxonomy, life cycle, and auxosporulation of Nitzschia fonticola (Bacillariophyta). Journal of Phycology. 42: 1353-1372. | Morphology; Taxonomy; Biology |
| Mann D.G., Trobajo R. 2014. Symmetry and sex in Bacillariaceae (Bacillariophyta), with descriptions of three new Nitzschia species. European Journal of Phycology. 49(3): 276-297. | Morphology; Biology; Illustrations |
| Mann D.G., Trobajo R., Sato S., Li C., Witkowski A., Rimet F., Ashworth M.P., Hollands R.M., Theriot E.C. 2021. Ripe for reassessment: A synthesis of available molecular data for the speciose diatom family Bacillariaceae. Molecular Phylogenetics and Evolution. 158: 106985. | Biology; Illustrations; Taxonomy; |
This page should be cited as:
Mann D. G. Nitzschia A.H. Hassall; 1845; 435. In: Jüttner I., Carter C., Cox E.J., Ector L., Jones V., Kelly M.G., Kennedy B., Mann D.G., Turner J. A., Van de Vijver B., Wetzel C.E., Williams D.M..
Freshwater Diatom Flora of Britain and Ireland. Amgueddfa Cymru - National Museum Wales. Available online at https://naturalhistory.museumwales.ac.uk/diatoms/browsespecies.php?-recid=3725. [Accessed:
].
Image Gallery






Record last modified: 27/12/2020