
Surirella P.J.F. Turpin; 1828; 362-363
Key references
Round F.E., Crawford R.M., Mann D.G. 1990. The Diatoms. Biology & Morphology of the Genera. Cambridge University Press, Cambridge. 747 pp
English J., Potapova M. 2012. Ontogenetic and interspecific valve shape variation in the Pinnatae group of the genus Surirella and the description of S. lacrimula sp. nov. Diatom Research. 27(1-2): 9-27.
Morphology
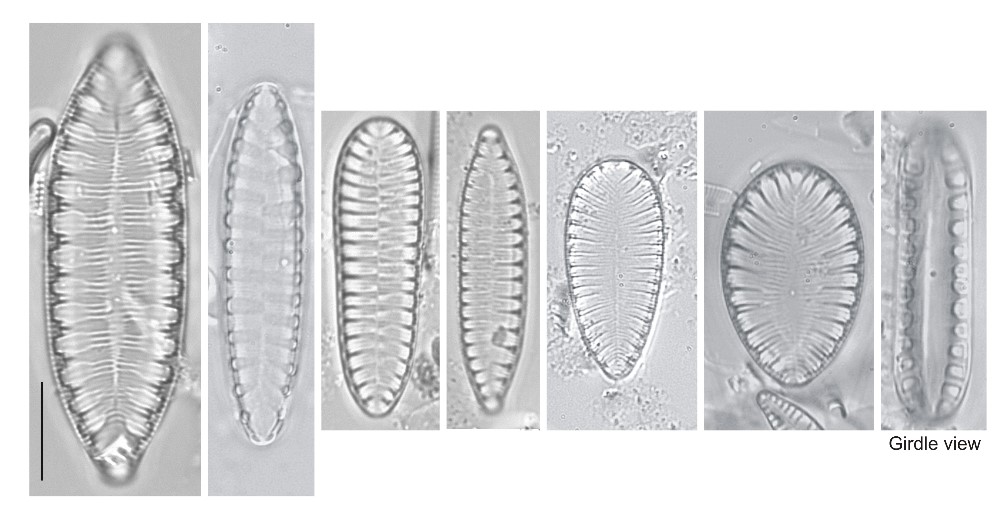
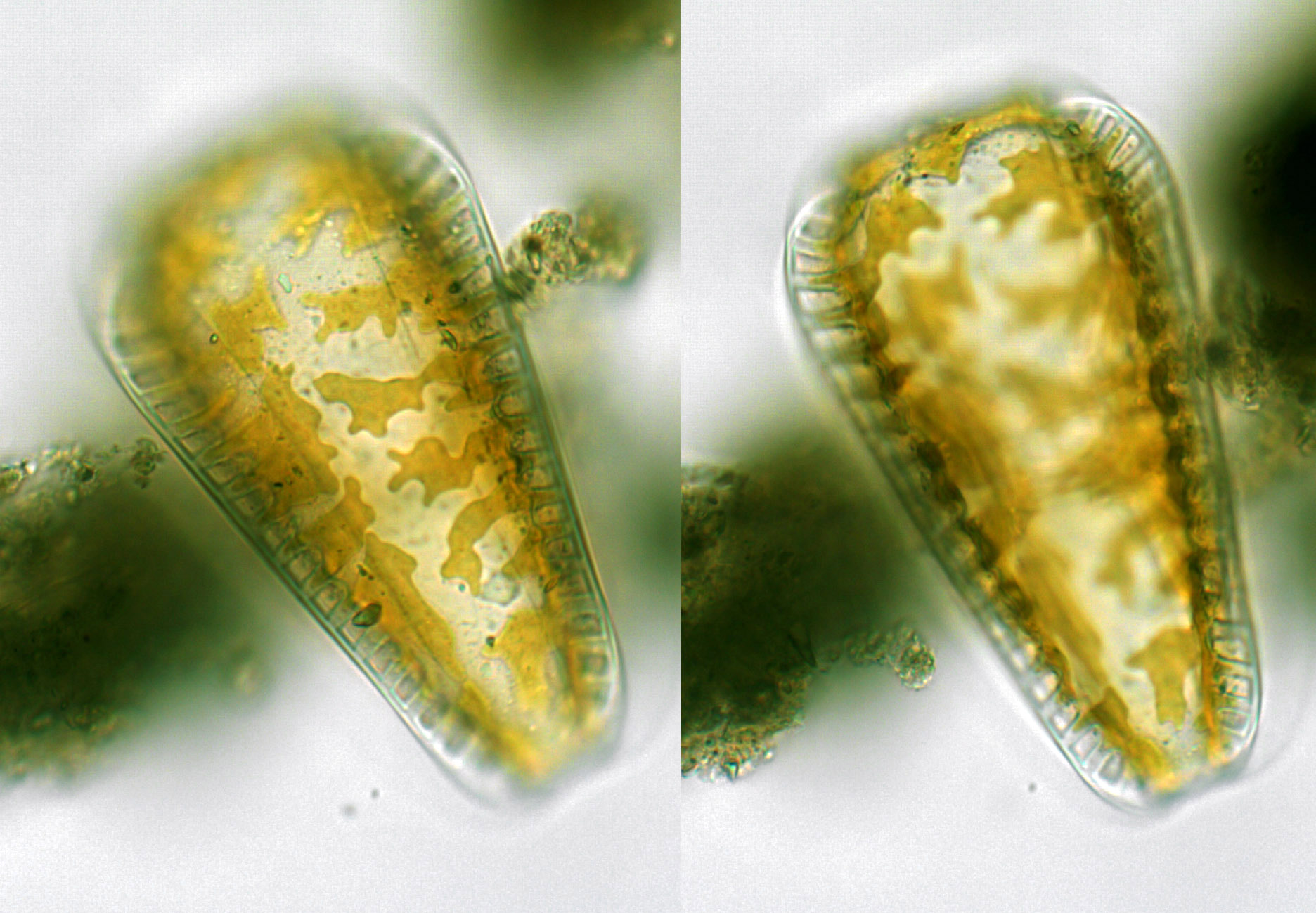
Cells seen in valve view or girdle view, depending on the species; isolated valves usually lying in valve view. Occasionally cells are spiralled. Valves bipolar in structure, with a definite longitudinal axis, but may be elliptical or even circular in shape; more often, however, valves are linear, lanceolate or rhombic.
Frustules isopolar or heteropolar, valves bilaterally symmetrical.
The valve pores are usually very small and the striae that they form are also sometimes inconspicuous. Much more obvious are rib-like thickenings and wavy infoldings of the valve, the areas of pores and striae that these surround, and the fibulae.
Axial area extremely narrow, indistinguishable in LM.
Raphe running around the whole perimeter of the valve and opening inwardly into a tubular canal ('canal-raphe') raised on a ridge or keel. The canal is separated from the remainder of the cell interior by siliceous bridges (fibulae) and often also by infoldings of the valve face and mantle, which, in some species where the keel is especially high, fuse to create extracellular windows (fenestrae) beneath the raphe. In LM, the raphe itself is almost impossible to detect: its presence must be deduced from the presence of the fibulae or fenestrae. Within a frustule, the raphe endings of the two valves are not offset as in Campylodiscus. One set of raphe endings is apparently equivalent to the central raphe endings of other raphid diatoms; the other set is apparently equivalent to the polar raphe endings, brought next to each other by reflexing of the valve poles in dorsiventral forms perhaps resembling extant Auricula.
One chloroplast per cell, consisting of two large plates, often lobed at their margins, connected by a narrow isthmus that lies at one end of the cell, or close to the nucleus in a subcentral position. The plates lie against the valves and, when seen in valve view, appear to fill all except the cell periphery. Along the longitudinal axis of the valve there is sometimes a median rib or hyaline area. This is only superficially similar to the sternum or raphe-sternum of other pennate diatoms. Valve development in Surirellales, as in other raphid diatoms except Eunotiales, occurs from the raphe system outwards.
Frustules isopolar or heteropolar, valves bilaterally symmetrical.
The valve pores are usually very small and the striae that they form are also sometimes inconspicuous. Much more obvious are rib-like thickenings and wavy infoldings of the valve, the areas of pores and striae that these surround, and the fibulae.
Axial area extremely narrow, indistinguishable in LM.
Raphe running around the whole perimeter of the valve and opening inwardly into a tubular canal ('canal-raphe') raised on a ridge or keel. The canal is separated from the remainder of the cell interior by siliceous bridges (fibulae) and often also by infoldings of the valve face and mantle, which, in some species where the keel is especially high, fuse to create extracellular windows (fenestrae) beneath the raphe. In LM, the raphe itself is almost impossible to detect: its presence must be deduced from the presence of the fibulae or fenestrae. Within a frustule, the raphe endings of the two valves are not offset as in Campylodiscus. One set of raphe endings is apparently equivalent to the central raphe endings of other raphid diatoms; the other set is apparently equivalent to the polar raphe endings, brought next to each other by reflexing of the valve poles in dorsiventral forms perhaps resembling extant Auricula.
One chloroplast per cell, consisting of two large plates, often lobed at their margins, connected by a narrow isthmus that lies at one end of the cell, or close to the nucleus in a subcentral position. The plates lie against the valves and, when seen in valve view, appear to fill all except the cell periphery. Along the longitudinal axis of the valve there is sometimes a median rib or hyaline area. This is only superficially similar to the sternum or raphe-sternum of other pennate diatoms. Valve development in Surirellales, as in other raphid diatoms except Eunotiales, occurs from the raphe system outwards.
Literature
References are given in chronological order.
Reference |
Citation |
|---|---|
| Turpin, P.J.F. 1828 ref 893. Observations sur le nouveau genre Surirella. Mémoires du Museum d'Histoire Naturelle. Paris, Vol:16, 361-368, pl. 15, 18 figs. | Morphology; Taxonomy; Description |
| Paddock T.B.B. 1978. Observations on the valve structure of diatoms of the genus Plagiodiscus and on some associated species of Surirella. Botanical Journal of the Linnean Society. 76: 1-25. | Morphology; Illustrations |
| Paddock T.B.B. 1985. Observations and comments on the diatom Surirella fastuosa and Campylodiscus fastuosus and other species of similar appearance. Nova Hedwigia. 41: 417-444. | Morphology; Illustrations |
| Krammer K., Lange-Bertalot H. 1987. Morphology and taxonomy of Surirella ovalis and related taxa. Diatom Research. 2(1): 77-95. | Morphology; Taxonomy; Illustrations |
| Round F.E., Crawford R.M., Mann D.G. 1990. The Diatoms. Biology & Morphology of the Genera. Cambridge University Press, Cambridge. 747 pp | Morphology; Illustrations; |
| Mann D.G. 1996. Auxospore formation and neoteny in Surirella angusta (Bacillariophyta) and a modified terminology for cells of Surirellaceae. Nova Hedwigia. 71: 165-183. | Biology; Illustrations; Morphology |
| Ruck E.C., Kociolek J.P. 2005. Preliminary phylogeny of the Family Surirellaceae (Bacillariophyta). 50. Bibliotheca Diatomologica. 1-236 pp | Biology |
| Bramburger A.J., Haffner G.D., Hamilton P.B., Hinz F., Hehanussa P.E. 2006. An examination of species within the genus Surirella from the Malili Lakes, Sulawesi Island, Indonesia, with descriptions of 11 new taxa. Diatom Research. 21(1): 1-56. | Morphology; Illustrations; Taxonomy |
| English J., Potapova M. 2012. Ontogenetic and interspecific valve shape variation in the Pinnatae group of the genus Surirella and the description of S. lacrimula sp. nov. Diatom Research. 27(1-2): 9-27. | ; Illustrations; Morphology |
| Van de Vijver B., Cocquyt C., de Haan M., Kopalová K., Zidarova R. 2012. The genus Surirella (Bacillariophyta) in the sub-Antarctic and maritime Antarctic region. Diatom Research. 28: 92-108. | Morphology; Illustrations; Taxonomy |
| Karthick B., Hamilton P.B., Kociolek J.P. 2012. Taxonomy and biogeography of some Surirella Turpin (Bacillariophyceae) taxa from Peninsular India. Nova Hedwigia, Beiheft. 141: 81-116. | Morphology; Taxonomy; Illustrations |
| Ruck E.C., Nakov T., Alverson A.J., Theriot E.C. 2016. Phylogeny, ecology, morphological evolution, and reclassification of the diatom orders Surirellales and Rhopalodiales. Molecular Phylogenetics and Evolution. 103: 155-171. | Morphology; Biology; Illustrations; Taxonomy |
| Ruck E.C., Nakov T., Alverson A.J., Theriot E.C. 2016. Nomenclatural transfers associated with the phylogenetic reclassification of the Surirellales and Rhopalodiales. Notularum algarum. No. 10: 1-4. | Taxonomy |
| Jahn R., Kusber W.-H., Cocquyt C. 2017. Differentiating Iconella from Surirella (Bacillariophyceae): typifying four Ehrenberg names and a preliminary checklist of the African taxa. PhytoKeys. 82: 73-112. | Taxonomy; Illustrations; Morphology |
This page should be cited as:
Mann D. G. Surirella P.J.F. Turpin; 1828; 362-363. In: Jüttner I., Carter C., Cox E.J., Ector L., Jones V., Kelly M.G., Kennedy B., Mann D.G., Turner J. A., Van de Vijver B., Wetzel C.E., Williams D.M..
Freshwater Diatom Flora of Britain and Ireland. Amgueddfa Cymru - National Museum Wales. Available online at https://naturalhistory.museumwales.ac.uk/diatoms/browsespecies.php?-recid=3746. [Accessed:
].
Image Gallery


Record last modified: 27/12/2020